Research
Research
Head of department:
RNDr. Karel Janko, Ph.D.
 |
|
Laboratory video
Download link: video.
 „Two things that are certain in life are death and taxes (Benjamin Franklin 1789) but before either death or taxes, generally comes fertilization…”
„Two things that are certain in life are death and taxes (Benjamin Franklin 1789) but before either death or taxes, generally comes fertilization…”
(Palumbi 2009).
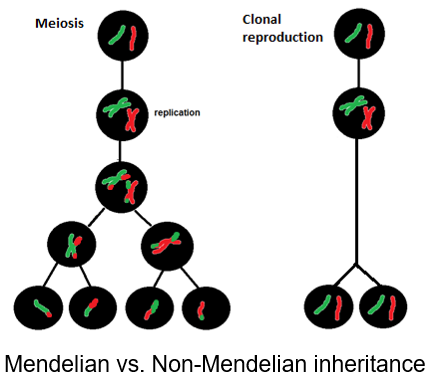 Broadly speaking, reproduction, the ability to transmit genomes from one generation to another, is a fundamental property of all living organisms. Such a transmission may be realized via vegetative propagation, but gametic reproduction is omnipresent. There are, however, a number of variations on this theme, ranging from sexual reproduction, with segregation of homologous chromosomes and recombination between them, to fully clonal, parthenogenetic, reproduction. Interestingly, asexual modes of reproduction are often associated with interspecific hybridization. We investigate how such changes in reproductive mode impact on genome evolution, phenotypes and, ultimately, the formation of new species. Our particular focus is to understand how cis- and trans- regulatory interactions between genes and subgenomes in hybrids relate to tier reproduction and performance.
Broadly speaking, reproduction, the ability to transmit genomes from one generation to another, is a fundamental property of all living organisms. Such a transmission may be realized via vegetative propagation, but gametic reproduction is omnipresent. There are, however, a number of variations on this theme, ranging from sexual reproduction, with segregation of homologous chromosomes and recombination between them, to fully clonal, parthenogenetic, reproduction. Interestingly, asexual modes of reproduction are often associated with interspecific hybridization. We investigate how such changes in reproductive mode impact on genome evolution, phenotypes and, ultimately, the formation of new species. Our particular focus is to understand how cis- and trans- regulatory interactions between genes and subgenomes in hybrids relate to tier reproduction and performance. - Frequent association of asexually reproducing organisms with ‘extreme’ environment at high latitudes and altitudes also brought us to investigate how Metazoans adapt to cold climatic conditions of polar regions and glaciers, in particular.
Model organisms
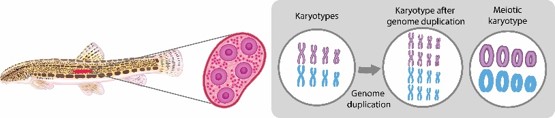
European spined loaches (Cobitis), include several phylogenetic lineages that independently colonized European drainages. They hybridize, producing several hybrid biotypes, comprising diploids, triploids and tetraploids. Hybrid females and males notably differ in their ability to reproduce; while diploid and triploid hybrid males are always sterile, hybrid females maintain fertility and reproduce via gynogenesis (=sperm-dependent parthenogenesis). Formation of unreduced eggs relies on premeiotic genome endoreplication, ensuring chromosome pairing in meiosis. This suggests genetic sex determination is linked to gametogenesis and reproductive modes.
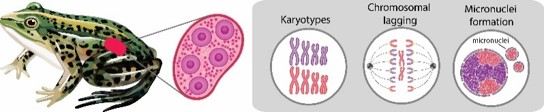
The Pelophylax esculentus complex includes two parental species, P. ridibundus and P. lessonae, as well as several forms of their interspecific hybrids P. esculentus. Hybrid frogs exist as diploid hybridogens and triploids reproducing by meiotic hybridogenesis. In contrast to other “asexual” vertebrates, P. esculentus is represented not only by females but also males. During the gametogenesis, one genome is retained and the other is excluded from the germ cells prior to meiosis. The genome elimination is gradual, proceeding during gonocyte multiplication and encapsulated in micronuclei, possibly by chromosomal lagging.
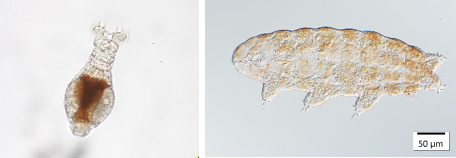
The phylum Tardigrada (also known as water bears) is famous for their extraordinary resistance to extremely high and low temperatures, over-drying, high doses of ionizing radiation and their anhydrobiotic state allowing them to survive even in the open-space. With more than 1300 species described worldwide, water bears live in a wide variety of aquatic and terrestrial environments and in some polar ecosystems such as nunataks, coastal plains or glaciers.
Europe: Czech republic, Poland, Balkan, Norway
Africa: Uganda
Asia: South Korea
America: Oklahoma
Antarctica
High Arctic
I am always on the lookout for bright and motivated collaborators. If you are interested in what we do or want to join us as a student or postdoc, please e-mail Karel Janko at janko@iapg.cas.cz and include your CV.
Call for a POSTDOC position open in the frame of MERIT program https://meritcb.eu/; https://www.youtube.com/watch?v=913tJvX64RA
Emergence of alternative reproductive modes:
- Most metazoans produce reduced and recombinant gametes for sexual reproduction, but some strains have abandoned traditional sexual reproduction and switched to asexual reproduction. Interestingly, many of these asexual lineages are of hybrid origin, suggesting that the aberrations in the meiotic cycle towards clonality are linked to impaired regulation of gametogenic programs in hybrids.
- We conduct studies to determine whether the emergence of asexual reproduction is correlated with genetic and chromosomal divergence among parental species. Our findings suggest that hybridization between closely related or similar species typically produces sexual hybrids, while increasing genetic divergence promotes the formation of asexual lineages. However, asexual development only accounts for a minor fraction of the hybrid's gametes, with the majority of its cells attempting to proceed through normal meiosis.
- Therefore, for a hybrid to form a successful clone, it must not only involve aberrations in gametogenic programs to initiate clonality but also a mechanism to halt the development of normal gametes, which can be achieved through aberrant pairing of chromosomes in hybrids between karyotypically divergent parents.
- Dedukh D, Majtánová Z, Marta A, Pšenička M, Kotusz J, Klíma J, Juchno D, Boron A, Janko K. 2020. Parthenogenesis as a Solution to Hybrid Sterility: the Mechanistic Basis of Meiotic Distortions in Clonal and Sterile Hybrids. Genetics 215:975–987; Dedukh D, Marta A, Janko K. 2021. Challenges and Costs of Asexuality: Variation in Premeiotic Genome Duplication in Gynogenetic Hybrids from Cobitis taenia Complex. International Journal of Molecular Sciences 22:12117).
Asexuality and speciation:
- The data thus points to the fact that asexuality in hybrids originates as a by-product of accumulated incompatibilities between parental genomes. Because asexuals may not effectively transmit genes between parental species, it may therefore be considered as a special type of Bateson-Dobzhansky-Muller incompatibility that originates hand-in-and with hybrid sterility.
- Janko K, Pačes J, Wilkinson-Herbots H, Costa RJ, Roslein J, Drozd P, Iakovenko N, Rídl J, Hroudová M, Kočí J, et al. 2018. Hybrid asexuality as a primary postzygotic barrier between nascent species: On the interconnection between asexuality, hybridization and speciation. Mol Ecol:249–263.; Stöck M, Dedukh D, Reifová R, Lamatsch DK, Starostová Z, Janko K. 2021. Sex chromosomes in meiotic, hemiclonal, clonal and polyploid hybrid vertebrates: along the “extended speciation continuum.” Philos Trans R Soc Lond B Biol Sci 376:20200103.
Evolution of asexual organisms:
- According to theory, the emergence of asexuality is a rare event, and once it occurs, asexual organisms should have a short lifespan due to the rapid accumulation of harmful mutations. Additionally, asexual organisms may struggle to adapt to changing environments and compete with other organisms and pathogens.
- However, our data shows that asexuality can be triggered relatively easily in some groups, and natural clones are surprisingly resistant to parasites and do not accumulate harmful mutations at high rates. The short-livedness of most natural asexuals thus results from their stochastic replacement by other clonal lineages via a drift-like process rather than deterministic processes such as Muller's ratchet.
- In summary, while theory predicts that asexuality is a rare and short-lived occurrence, our findings suggest that natural asexuals can persist for extended periods and resist parasites and mutations. This highlights the importance of investigating the mechanisms that allow for the persistence and success of asexual organisms in the natural world.
- Janko K, Drozd P, Eisner J. 2011. Do clones degenerate over time? Explaining the genetic variability of asexuals through population genetic models. Biology Direct 6:17.; Janko K, Drozd P, Flegr J, Pannell JR. 2008. Clonal turnover versus clonal decay: a null model for observed patterns of asexual longevity, diversity and distribution. Evolution 62:1264–1270.; Kočí J, Röslein J, Pačes J, Kotusz J, Halačka K, Koščo J, Fedorčák J, Iakovenko N, Janko K. 2020. No evidence for accumulation of deleterious mutations and fitness degradation in clonal fish hybrids: Abandoning sex without regrets. Molecular Ecology 29:3038–3055.; Kotusz J, Popiołek M, Drozd P, De Gelas K, Šlechtová V, Janko K. 2014. Role of parasite load and differential habitat preferences in maintaining the coexistence of sexual and asexual competitors in fish of the Cobitis taenia hybrid complex. Biol J Linn Soc Lond 113:220–235.

Genomics and transcriptomics of speciation:
- When two species interbreed, their distinct regulatory networks are combined within the same cellular environment. Our team is investigating how this mixture affects the hybrid's genetic, transcriptomic, and physical characteristics. We use asexual hybrids as models for our research because they maintain fixed heterozygosity, similar to the F1 generation, for many generations.
- Through a multidisciplinary approach, we have discovered that asexual genomes remain static on a large scale, particularly when examining entire chromosomes. However, at the individual gene level, they are constantly evolving due to changes in cis- and trans-regulatory interactions between subgenomes, as well as gene conversions and hemizygous deletions in specific pathways. This phenomenon can lead to "adaptive gene loss," where genes that are no longer necessary are deleted to streamline the hybrid's gene expression.
- In summary, our research focuses on how hybridization can result in dynamic genetic changes, particularly at the individual gene level. The focus on asexual hybrids provides insights into the evolution of hybrid genomes and the regulatory interactions that shape their phenotypic traits.
- Bartoš O, Röslein J, Kotusz J, Paces J, Pekárik L, Petrtýl M, Halačka K, Štefková Kašparová E, Mendel J, Boroń A, et al. 2019. The Legacy of Sexual Ancestors in Phenotypic Variability, Gene Expression, and Homoeolog Regulation of Asexual Hybrids and Polyploids. Mol Biol Evol 36:1902–1920.; Janko K, Bartoš O, Kočí J, Roslein J, Drdová EJ, Kotusz J, Eisner J, Mokrejš M, Štefková-Kašparová E. 2021. Genome Fractionation and Loss of Heterozygosity in Hybrids and Polyploids: Mechanisms, Consequences for Selection, and Link to Gene Function. Molecular Biology and Evolution 38:5255–5274.; Majtánová Z, Choleva L, Symonová R, Ráb P, Kotusz J, Pekárik L, Janko K. 2016. Asexual Reproduction Does Not Apparently Increase the Rate of Chromosomal Evolution: Karyotype Stability in Diploid and Triploid Clonal Hybrid Fish (Cobitis, Cypriniformes, Teleostei). PLoS ONE 11:e0146872.
Polar ecology and adaptation to extreme environments
- We currently investigate the diversity, speciation and population genetic patterns of soil and glacial tardigrades, rotifers and nematodes inhabiting polar regions. Our main interest focuses on understanding how these creatures move between regions and isolated ice patches and whether they have phylogeographic patterns at all? Why are they so endemic and to what extent are their distributions driven by geography as opposed to habitat selection.
- In previous projects, we have investigated adaptive radiations of Notothenioid fish in relation to the massive cooling Antarctic continent and the Southern Ocean. This work was chilly and cool at the same time and revealed how the tempo of radiation correlated with climatic changes.
- Furthermore, we compared the phylogeographic patterns between benthic and pelagic fish species and discovered that climatic cycles had a significantly greater impact on the former group. This finding implies that the primary impact of climate change was not solely due to changes in temperature or climate, but rather due to the destruction of habitats caused by advancing and retreating glaciers.
- Iakovenko NS, Smykla J, Convey P, Kašparová E, Kozeretska IA, Trokhymets V, Dykyy I, Plewka M, Devetter M, Duriš Z, et al. 2015. Antarctic bdelloid rotifers: diversity, endemism and evolution. Hydrobiologia 761:5–43.; Zawierucha K, Buda J, Jaromerska TN, Janko K, Gąsiorek P. 2020. Integrative approach reveals new species of water bears (Pilatobius, Grevenius, and Acutuncus) from Arctic cryoconite holes, with the discovery of hidden lineages of Hypsibius. Zoologischer Anzeiger 289:141–165.; Zawierucha K, Marshall CJ, Wharton D, Janko K. 2019. A nematode in the mist: Scottnema lindsayae is the only soil metazoan in remote Antarctic deserts, at greater densities with altitude. Polar Research.; Janko K, Marshall C, Musilová Z, Houdt JV, Couloux A, Cruaud C, Lecointre G. 2011. Multilocus analyses of an Antarctic fish species flock (Teleostei, Notothenioidei, Trematominae): Phylogenetic approach and test of the early-radiation event. Molecular Phylogenetics and Evolution.; Janko K, Lecointre G, DeVries A, Couloux A, Cruaud C, Marshall C. 2007. Did glacial advances during the Pleistocene influence differently the demographic histories of benthic and pelagic Antarctic shelf fishes? – Inferences from intraspecific mitochondrial and nuclear DNA sequence diversity. BMC evolutionary biology 7:220.; Kašparová E, Putte APV de, Marshall C, Janko K. 2015. Lifestyle and Ice: The Relationship between Ecological Specialization and Response to Pleistocene Climate Change. PLOS ONE 10:e0138766

We collaborate on extremophilous communities of glacial invertebrates.with groups of:
- Krszysztof Zawirerucha (http://ztez.amu.edu.pl/en/staff/assistant-professors/122-krzysztof-zawierucha-m-sc)
- Diego Fontaneto (https://publications.cnr.it/authors/diego.fontaneto)
- Dag Hessen (https://www.mn.uio.no/ibv/english/people/aca/hessen/index.html).
- Kamil Jaron (https://www.sanger.ac.uk/group/jaron-group/)
We have common projects on ecology, evolution and genomics of asexual organisms with groups of:
- Matthias Stock (https://www.igb-berlin.de/en/profile/matthias-stock)
- Ingo Schlupp (https://www.ou.edu/cas/biology/people/faculty/ingo-schlupp)
- Yong Jin Won (https://pure.ewha.ac.kr/en/persons/yong-jin-won)
- Jan Kotusz (http://www.muzeum-przyrodnicze.uni.wroc.pl/php?go=badania-naukowe-kotusz)
- Aurora Ruiz Herrera (https://webs.uab.cat/evolgenom/staff/pi/)